-
2025-04-25 06:55:59
РЕПОРТЕР ЭСЭД ЭНДОПЛАЗМЫН ТОРЛОГИЙН СТРЕССИЙГ ЦЕЛАСТРОЛООР ЗОХИЦУУЛАХ НӨЛӨӨГ СУДАЛСАН ДҮН
РЕПОРТЕР ЭСЭД ЭНДОПЛАЗМЫН ТОРЛОГИЙН СТРЕССИЙГ ЦЕЛАСТРОЛООР ЗОХИЦУУЛАХ НӨЛӨӨГ СУДАЛСАН ДҮН
Үндэслэл
Целастрол нь Celastraceae овгийн Tripterygium wilfordii Hook F (TWHF)-ийн үндэсний хальснаас гаргаж авсан тритерпенойд бөгөөд олон төрлийн фармакологийн үйлчилгээтэй нэгдэл юм. Целастрол нь хоруу чанар багатай, өргөн цар хүрээний олон төрлийн үйлчилгээтэй гэх мэт шинж чанараараа олон төрлийн хорт хавдрын эсрэг эмчилгээний идэвхит бодис болохын хувьд сүүлийн жилүүдэд анхаарал татаж байна1.
Гадаад болон дотоод хүчин зүйлүүдийн нөлөөгөөр эсийн эндоплазмын торын хөндий дотор уураг орон зайд дутуу эсвэл алдаатай нугаларан хуримтлагдаж эхэлснээр эс стресст өртдөг2. Энэ стресст өртсөн эс эндоплазмын торлогийн гомеостазыг сэргээх зорилгоор хариу үйлдэл (unfolded protein response (UPR)) үзүүлдэг. Энэ хариу үйлдлийн үед эсийн уургийн орон зайд нугаларах зохицуулга эрчимжих, уургийн нийлэгжил түр зуур удаашрах, алдаатай нугаларсан уургууд болон гэмтсэн эрхтэнцрүүд задарч устах явц эрчимжинэ3. Нугалаараагүй буюу алдаатай нугаларсан уургууд нь ЭТ-д байрладаг уураг (BiP (GRP78))–тай холбогдож, UPR дохиоллыг дамжуулдаг IRE1α/XBP1, PERK/eIF2α/ATF4/CHOP ба ATF6α/ CHOP салбар замуудыг идэвхижүүлдэг4.
HLR-GV-ATF6α (цаашид репортер эс гэх) нь БНСУ-ын ЧҮИС-ийн Био-Анагаахын тэнхимийн лабораторид гаргаж авсан рекомбинант HeLa эс бөгөөд ЭТ стрессийн үед ATF6α уургийн генийн транскрипцийн идэвхижлийг хянадаг репортер эс юм. Энэ репортер эс нь ~ 70 кДа молекул жинтэй GV-hATF6αΔN (a.х 333-670) рекомбинант (fusion) уургийг ЭТ-ийн мемран дээр байрладаг трансмембран уураг хэлбэрээр экпрессэлдэг. Эс ЭТ стрессэд өртсөн нөхцөлд дээрх нэгдмэл уураг нь ЭТ-оос Гольджийн аппарат руу шилжин байршиж, протеаза энзимээр хэрчигдэнэ. Үүссэн ~40 кДа молекул жинтэй GV hATF6αΔNΔC (a.х 333-380) фрагмент нь эсийн бөөм рүү шилжсэнээр ЭТ стрессийн хариу үйлдлийн үйл явцад транскрипцийн факторын үүрэг гүйцэтгэдэг6. Тапсигаргин нь (Тг) сарко-/эндоплазмын торлогийн Са+2 АТФ(SERCA)-азын ингибитор бөгөөд ЭТ стресс болон UPR-ын биологийг судлах сонгодог хэрэгсэл юм7. Одоогоор энэ репортер эс дээр ЭТ стрессийн үед ЭТ стресс болон апоптозтой холбоотой биомаркер уургуудын экспрессийн нарийвчилсан судалгаа хийгдээгүй байна.
Зорилго
Энэ судалгаагаар бид тапсигаргинаар өдөөгдсөн ЭТ стрессийн үед репортер эсийн хариу үйлдэл болон түүнд целастрол хэрхэн нөлөөлөхийг судлах зорилго тавив.
Материал, арга зүй
Эсийн өсгөвөр: 5х104 эс/мл HLR-GV-ATF6α репортер эсийг 10% ийн үхрийн хээлийн ийлдэс, 1%-ийн антибиотикийн холимог, 1X глутамакс, 0.1 мг/мл хигромицин B and 0.1 мг/мл генетицин бүхий DMEM орчинд 5% СО2 инкубаторт 37°С хэмд 24 цаг өсгөвөрлөөд, өгөгдсөн концентрацитай Тг болон целастрол бүхий орчинд өгөгдсөн цагийн турш үргэлжүүлэн өсгөвөрлөж шинжилгээг хийсэн.
Эсийн амьдрах чадвар: Эсийн амьдрах чадварыг МТТ колориметрийн аргаар шинжилсэн. 96 үүрт эсийн өсгөврийн хавтанд репортер эсийг өсгөвөрлөж, өгөгдсөн концентраци бүхий Тг болон целастролын хамт 48 цаг инкубаци хийсний дараа, 10 мкл MTT уусмалыг нэмж 4 цаг СО2 инкубаторт 37°С т байлгана. Эсийн өсгөвөрийн шингэнийг соруулж аваад формазан талстыг 100 мкл DMSO нэмж уусгана. Эсийн өсгөвөрт тиазолил хөх тэтразолын бромидийн өнгө хувирлыг 540 нм-т бичил хавтан уншигч дээр хэмжсэн. Эсийн амьдрах чадварыг хяналтын бүлгийн хувиар илэрхийлсэн. Тг-ыг диметилсульфоксидод (ДМСО) уусгасан тул хяналтын уусмалаар ДМСО-ыг хэрэглэсэн.
Люциферазийн идэвхжил: Репортер эсийг өгөгдсөн концентраци бүхий Тг болон целастролын хамт 12 цаг өсгөвөрлөж, ATF6α-ийн идэвхижлийг люциферазийн сорилын системийг (Promega) ашиглан холбогдох зааврын дагуу 20 мкл эсийн лизатанд шинжилсэн. Люциферазийн идэвхийг эсийн лизат бүрт 1 мкг уургийн люцифераз идэвхийн абсолют хэмжээгээр үзүүлсэн.
Вестерн блот: Өгөгдсөн концентрац бүхий целастрол, Тг болон Тг/целастролын хамт 6, 12, 24 and 48 цагийн турш тус тус өсгөвөрлөөд уургийн экспрессийн түвшинг вестерн блотын аргаар шинжилж, β-актин уургийг эерэг хяналт болгон хэрэглэсэн. Өсгөвөрлөсөн эсээс цитоплазмын ба бөөмийн уургийг ялгах кит ашиглан холбогдох зааврын дагуу ялгасны дараа 8-12% натрийн додецил сульфат полиакриламидын гель электрофорезоор (SDS-PAGE) салгаж, PVDF мембран руу шилжүүлсэн. Мембраныг 5% ийлдэсний альбумин эсвэл тосгүй сүү агуулсан 1xT (Tвин 20)-Трис буферээр блоколсны дараа, эхлээд GRP78, PERK, CHOP, caspase-3, PARP, p-eIF2α, β-актин, тюбулин, ламин А болон GAL4 DNA-BD зэрэг анхдагч эсрэг биеүдтэй, дараа нь үргэлжлүүлэн HRP-хулгана ба туулайн эсрэг IgG-ийн эсрэг биеийн коньюгаттай инкубаци хийсэн. Уурагтай холбогдсон эсрэг биеүдийг Amersham Imager 600 химилюминес-ценцийн систем дээр шинжилсэн.
Статистик шинжилгээ: Өгөгдлийг гурван тусдаа туршилтын ±SEM-ээр, хяналтын бүлэгтэй харьцуулан илэрхийлсэн. Вестерн блотын өгөгдлийг GraphPad Prism5 (GraphPad Software, Inc, La Jolla, CA, USA) программаар шинжлэн тооцоо хийсэн. Бүх өгөгдлийн тооцооллыг Microsoft Excel программ ашиглан хийсэн.
Үр дүн
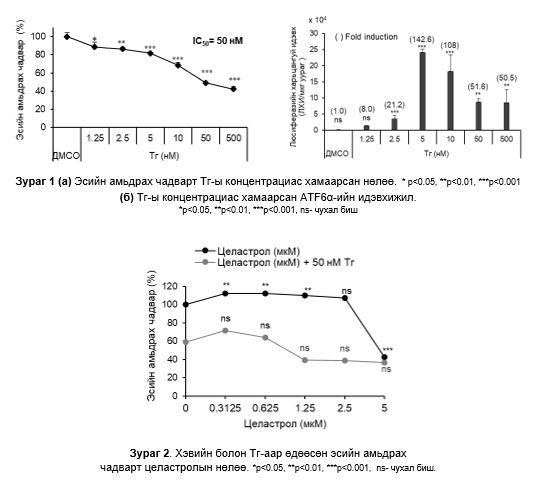
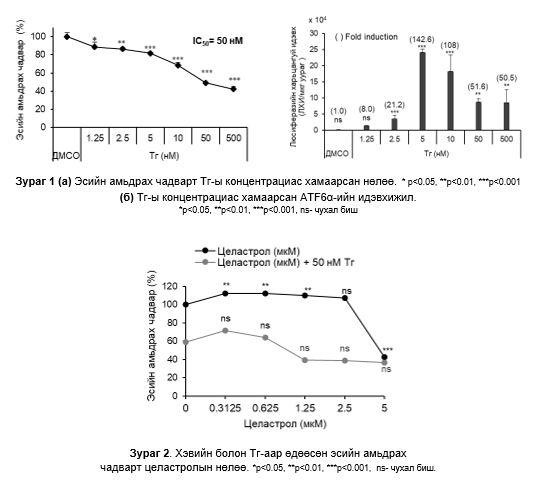
Эсийн амьдрах чадвар ба ATF6α-ын идэвхижилд Тг-ы нөлөө: Тг-ы концентраци ихсэх тусам эсийн амьдрах чадвар буурч байсан ба эсийн амьдрах чадварыг 50% -иар бууруулах Тг-ы концентраци нь 50 нM (IC50) байсан (Зураг 1a)
Люциферазийн хамгийн өндөр идэвхи Тг-ы 5 нM утганд илэрсэн. Тг-ы концентраци ихсэх тусам люциферазийн идэвх суларсан (Зураг 1б). Хаалтанд байгаа тоо нь хяналтын бүлэгтэй харьцуулсан дундаж өдөөлтийг илэрхийлсэн.
Эсийн амьдрах чадвар ба ATF6α-ийн идэвхижилд целастролын нөлөө: Репортер эсийг 48 цагийн турш 0.3125-5 мкМ целастролтай хамт өсгөвөрлөхөд эсийн амьдрах чадвар 5 мкМ целастролын утганд 60%-иар буурсан. Харин 0.3125-5 мкМ целастрол болон 50 нM Тг-тай хамт өсгөвөрлөхөд эсийн амьдрах чадвар 60-70%-иар буурч болох хамгийн бага концентраци нь 1.25 мкM байсан тул дараачийн шинжилгээнд сонгон хэрэглэсэн (Зураг 2).
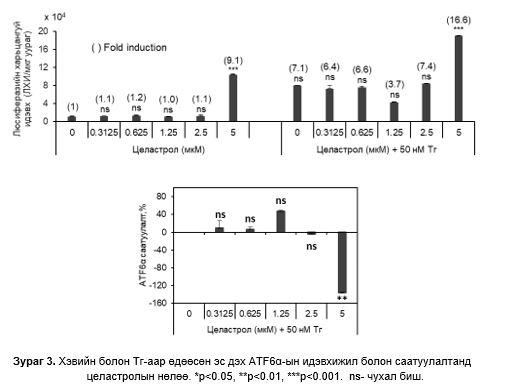
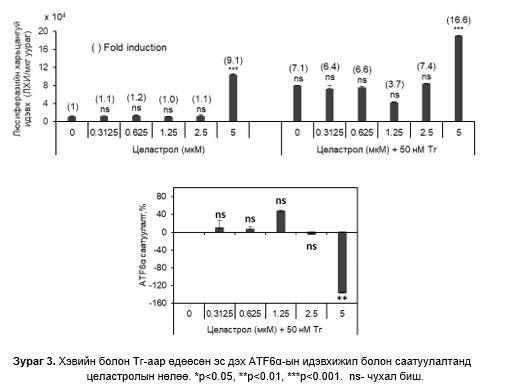
0-2.5 мкM целастрол бүхий өсгөвөрт люциферазийн идэвхи нэмэгдээгүй, 5 мкM (IC50) целастролын утганд 9.1х104 дахин өссөн. 50 нМ Тг ба 1.25 мкM целастролын утганд люциферазийн идэвхи хамгийн бага, ATF6α –идэвхижилийг саатуулалт хамгийн их байсан. 5 мкM- целастролын утганд люциферазийн идэвхи хамгийн өндөр, ATF6α идэвхижилийг саатуулалт хамгийн бага байсан (Зураг 3).
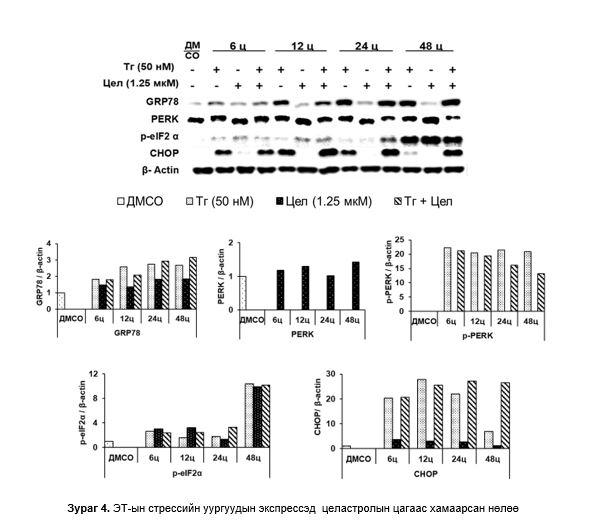
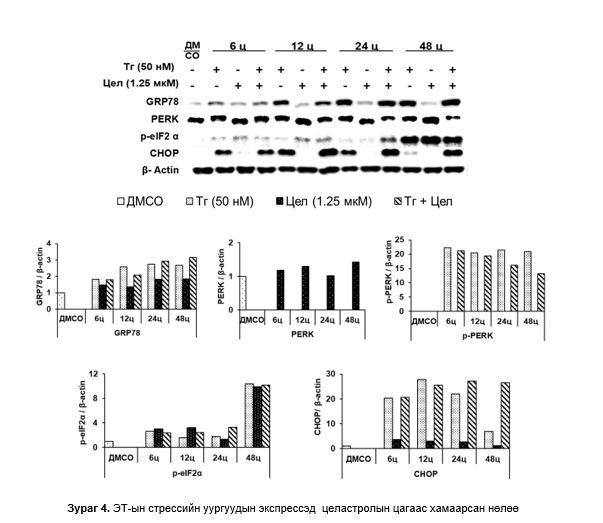
ЭТ-стрессийн уургуудын экспрессэд целастролын нөлөө: GRP78-ийн экспрессийн түвшин Тг-тай өсгөвөрт аажмаар өсөж 12-48 цагт жигд өндөр түвшинд байсан бол целастрол/Тг-тай өсгөвөрт 6-48 цагт түвшин нь өссөөр 48 цагт хамгийн өндөр байсан. Харин целастролтай өсгөвөрт 6-48 цагт аажмаар маш бага өссөн. PERK целастролтай өсгөвөрт мэдэгдэхүйц өсөж 48 цагт хамгийн өндөр байсан бол нөгөө 2 бүлэгт PERK илрээгүй, p-PERK Тг-тай өсгөвөрт 6-48 цагт хяналтын өсгөврөөс тогтмол өндөр байсан (уургийн зурвас мэдэгдэхүйц дээш шилжсэн). Целастрол/Тг-тай өсгөвөрт 6 цагт хамгийн өндөр байсан бол буурсаар 48 цагт хамгийн бага илэрсэн. Целастролтай өсгөвөрт p-PERK илрээгүй. p-eIF2α 3 бүлгийн өсгөвөрт 6-24 цагт бага хэмжээнд өсөөд, 48 цагийн дараа хамгийн өндөр байсан. CHOP Тг-тай өсгөвөрт эхний 12 цагт хамгийн дээд түвшинд хүрээд буурсаар 48 цагт хамгийн доод түвшинд хүрсэн бол целастрол/Тг-тай өсгөвөрт 12-24 цагт хамгийн өндөр хэвээр байсан. Целастролтай өсгөвөрт CHOP илрээгүй (Зураг 4).
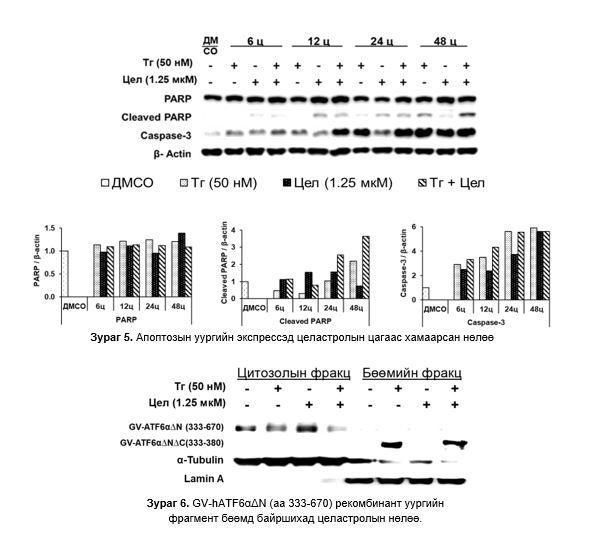
Апоптозын уургийн экспрессэд целастролын нөлөө: PARP дээрх 3 бүлэгт хяналттай харьцуулбал бага зэрэг өсөж ойролцоо түвшинд байсан гэхдээ целастролтай өсгөвөрт 48 цагт хамгийн өндөр илэрсэн. cl-PARP Тг- болон целастрол/Тг-тай өсгөвөрт 24 цагаас эхлэн өсөж, 48 цагт хамгийн өндөр бол целастролтай өсгөвөрт 12-24 цагт бага зэрэг өсөж 48 цагт буурсан. 3 бүлэгт Caspase-3 тогтмол нэмэгдсээр 48 цагт адилхан хамгийн өндөр түвшинд хүрсэн (Зураг 5).
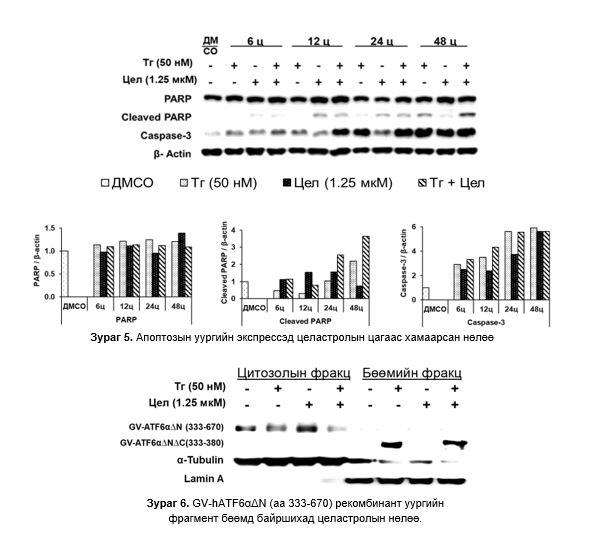
GV hATF6αΔNΔC (a.х 333-380) фрагмент бөөмд байршихад целастролын нөлөө: 70 кДа хэмжээтэй GV-hATF6αΔN (а.х. 333-670) уураг нь хяналтын бүлэг болон Тг, целастрол ба Тг/целастролтай өсгөвөрт эсийн цитоплазмын уургийн фракцад илэрсэн. ~40 кДа GV hATF6αΔNΔC (a.х 333-380) уураг нь Тг ба Тг/целастролтай өсгөвөрт бөөмийн уургийн фракцад илэрсэн. Целастрол бүхий өсгөвөрт ~40 кДа уураг илрээгүй (Зураг 6).
Хэлцэмж
Целастрол нь антиоксидант8, мэдрэлийг хамгаалах9, үрэвслийн эсрэг10, хорт хавдрын эсрэг11,12, остеоартритын эсрэг13,14, болон ангиогений эсрэг15 үйлчилгээтэй, хорт хавдрын эсэд апоптозыг өдөөдөг16 болох нь судалгаагаар тогтоогдсон.
Төрөл бүрийн хорт хавдрын явцыг удаашруулах, үр дүнтэй эмчлэхэд целастролын хорт хавдрын эсийн тархалт, шилжилт үсэрхийллийг саатуулах, мөн эсийн апоптозыг өдөөх, аутофаг болон ангиогенезийг дарангуйлах зэрэг олон молекул механизмууд нягт холбоотой нь батлагдсан. Мөн целастролын хавдрын эсрэг эмчилгээний гол бай болдог дохионы замуудыг судлан тогтоосон1.
ЭТ стрессийн үед GRP78-аас чөлөөлөгдсөн PERK сенсор уураг нь аутофосфоржих (p-PERK), eIF2α-ыг фосфоржуулах, ATF4-ын экспрессийг эрчимжүүлэх замаар, CHOP-ыг идэвхижүүлдэг. Хэрэв стресс нь зогсолтгүй, хүчтэй үргэлжилбэл ATF4-CHOPын идэвхижил нь апоптозыг үүсэх
эхлэлийг тавьдаг17,18. ATF6α нь идэвхижсэнээр CHOP-ын транскрипцийг, CHOP нь caspase-3-ын транскрипцийг тус тус идэвхижүүлж апоптозын үйл явц үргэлжилнэ19.
Бидний судалгаагаар, Тг-ы нөлөөгөөр эсэд ЭТ-ын стрессийн уургууд (GRP78, p-PERK, p-eIF2α ба CHOP) болон апоптозын уургуудын (Cl-PARP ба Caspase-3) экспрессийн түвшин өсөж байгаа нь эс стресст өртөж PERK болон ATF6α-ын зохицуулгын замууд идэвхижсэн төдийгүй апоптозын үйл явц эхэлснийг харуулж байна. Целастрол нь Tг-аар өдөөгдсөн эс дэх p-PERK-ийн экспрессийг бууруулсан боловч CHOP болон Cl-PARP-ийн экспрессийг нэмэгдүүлж байгаа нь целастрол Тг-тай хослон ЭТ стрессийг эрчимжүүлж байна.
~40 кДа уургийн фрагмент Tг болон целастрол/Tг бүхий өсгөврийн эсийн бөөмийн фракцад илэрч, хяналтын болон целастрол бүхий эсийн өсгөвөрт илрээгүй байгаа нь Тг-ы нөлөөгөөр ~40 кДа уураг цитоплазмын ~70 кДа уургаас тасарч бөөм рүү шилжсэнийг харуулж байна. Мөн энэ үйл явц ATF6α транскрипцийн фактор идэвхижсэнийг харуулж байна. 5 мкM целастрол нь хэвийн болон Tг-аар өдөөгдсөн эсүүдэд ATF6α идэвхижил өндөр байгаа нь энэ тунг ашиглан цаашид судлах хэрэгтэй гэж үзсэн.
Дүгнэлт
1. Целастрол p-PERK-ийн экспрессийг дарангуйлсан харин бусад ЭТ стресстэй холбоотой уургийн экспрессэд саатуулах нөлөө үзүүлээгүй. Целастрол Tг-тай хослон ЭТ стрессээс үүдэлтэй CHOP-ын экспресс болон PARP-ын задралыг идэвхижүүлж байгаа нь апоптозыг эрчимжүүлэх нөлөө үзүүлж байна.
2. Целастролыг хорт хавдрын эсрэг эмчилгээнд хэрэглэх зорилгоор түүний хавдрын эсэд ЭТ стресс болон апоптоз үүсгэх механизмыг уургийн дохиоллын түвшинд нарийвчлан судалж байгаагаараа энэ судалгаа шинэ эм хөгжүүлэх нийгэм эдийн засгийн болон шинжлэх ухааны ач холбогдолтой юм.
3. Энэ судалгаа нь Монгол орны эмийн ургамал, биологийн идэвхит бодисын үйлчлэлийн механизмын судалгаанд хэрэглэхэд хувь нэмэр болох ач холбогдолтой.
Ном зүй
1. C.Wang et al. Celastrol as an emerging anticancer agent: Current status, challenges and therapeutic strategies Biomedicine & Pharmacotherapy 163(2023)114882 https://doi.org/10.1016/j.biopha.2023.114882
2. Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science 2011; 334: 1081–1086.
3. Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature 2016; 529: 326–335.
4. Hetz C. Chevet E, Oakes SA. Proteostasis control by the unfolded protein response. Nat Cell Biol 2015; 17: 829–838.
5. Otero, J.H.; Lizák, B., Hendershot, L.M. Life and death of a bip substrate. Semin Cell Dev Biol. 2010, 21, 472–478.
6. Jin-Ik Kim, Randal J. Kaufman, Sung Hoon Back, and Ja-Young Moon. Development of a Reporter System Monitoring Regulated Intramembrane Proteolysis of the Transmembrane bZIP Transcription Factor ATF6α Mol Cells 2019; 42(11), 783-793
7. Jakobsen CM, Denmeade SR, Isaacs JT, Gady A, Olsen CE, Christensen SB. Design, synthesis, and pharmacological evaluation of thapsigargin analogues for targeting apoptosis to prostatic cancer cells. J Med Chem.2001;44(26):4696–4703. doi:10.1021/jm010985a
8. Trott, J.D. West, L. Klai_c, S.D. Westerheide, R.B. Silverman, R.I. Morimoto, K.A. Morano, Activation of heat shock and antioxidant responses by the natural product celastrol: transcriptional signatures of a thiol-targeted molecule, Mol. Biol. Cell 19 (2008) 1104-1112, https://doi.org/10.1091/ mbc e07-10-1004.
9. Cleren, N.Y. Calingasan, J. Chen, M.F. Beal, Celastrol protects against MPTPand 3-nitropropionic acid-induced neurotoxicity, J. Neurochem. 94 (2005) 995-1004,
https://doi.org/10.1111/j.1471-4159.2005.03253.x.
10. M. Hu, Q. Luo, G. Alitongbieke, S. Chong, C. Xu, L. Xie, X. Chen, D. Zhang, Y. Zhou, Z. Wang, X. Ye, L. Cai, F. Zhang, H. Chen, F. Jiang, H. Fang, S. Yang, J. Liu, M.T. Diaz-Meco, Y. Su, H. Zhou, J. Moscat, X. Lin, X. kun Zhang, Celastrol-inducedNur77 interaction with TRAF2 alleviates inflammation by promoting mitochondrial ubiquitination and autophagy, Mol. Cell 66(2017)141-153, https://doi.org/10.1016/j.molcel.2017.03.008, 6.
11. R. Kannaiyan, M.K. Shanmugam, G. Sethi, Molecular targets of celastrol derived from Thunder of God Vine: potential role in the treatment of inflammatory disorders and cancer, Cancer Lett. 303 (2011) 9-20, https:// doi.org/10.1016/j.canlet.2010.10.025.
12. D. Kashyap, A. Sharma, H.S. Tuli, K. Sak, T. Mukherjee, A. Bishayee, Molecular targets of celastrol in cancer: recent trends and advancements, Crit. Rev. Oncol. Hematol. 128 (2018) 70-81, https://doi.org/10.1016/ j.critrevonc.2018.05.019.
13. L. Gu, W. Bai, S. Li, Y. Zhang, Y. Han, Y. Gu, G. Meng, L. Xie, J. Wang, Y. Xiao, L. Shan, S. Zhou, L. Wei, A. Ferro, Y. Ji, Celastrol prevents atherosclerosis via inhibiting LOX-1 and oxidative stress, PLoS One 8 (2013) 1-11,
https:// doi.org/10.1371/journal.pone.0065477.
14. Q.H. Ding, Y. Cheng, W.P. Chen, H.M. Zhong, X.H. Wang, Celastrol, an inhibitor of heat shock protein 90b potently suppresses the expression of matrix metalloproteinases, inducible nitric oxide synthase and cyclooxygenase-2 in primary human osteoarthritic chondrocytes, Eur. J. Pharmacol. 708 (2013) 1-7, https://doi.org/10.1016/j.ejphar.2013.01.057
15. X.Pang, Z. Yi, J.Zhang, B. Lu, B. Sung, W. Qu, B.B. Aggarwal, M. Liu, Celastrol suppresses angiogenesis-mediated tumor growth through inhibition of AKT/ mammalian target of rapamycin pathway, Cancer Res. 70 (2010) 1951-1959, https://doi.org/10.1158/0008-5472.CAN-09-3201
16. Andrew M F, Justin R M, Amy L B, Danielle M G, Qinghua Z, Tyler E R, Neha N, Peter C, Yue Xi 2, Michael U Callaghan 2, Vamsi Kodali 7, Randal J Kaufman 7 Celastrol induces unfolded protein response-dependent cell death in head and neck cancer. Exp Cell Res. (2015) 412-422. DOI: 10.1016/j.yexcr.2014.08.014
17. Clarke H J., Chambers JE, Liniker E, Marciniak, S.J. Endoplasmic Reticulum Stress in Malignancy. Cancer Cell 2014, 25, 563–573.
18. Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004, 11, 381–389.
19. Han J, Back SH, Hur J, Lin YH, Gildersleeve R, Shan J, Yuan CL, Krokowski D, Wang S, Hatzoglou M, Kilberg MS, Sartor MA, Kaufman RJ. ER-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat Cell Biol. 2013; 15:481–490.
Ашигласан зураг